The Combination Effect of Gamma Irradiation and Salt Concentration on 2-Acetyl-1-Pyrroline Content, Proline Content and Growth of Thai Fragrant Rice (KDML 105)
Sompong Sansenya*1 , Yanling Hua2, Saowapa Chumanee3 and Chanun Sricheewin4
, Yanling Hua2, Saowapa Chumanee3 and Chanun Sricheewin4
1Department of Chemistry, Faculty of Science and Technology, Rajamangala University of Technology Thanyaburi, Pathum Thani 12110, Thailand.
2The Center for Scientific and Technological Equipment, Suranaree University of Technology, Nakhon Ratchasima 30000, Thailand.
3Division of Chemistry, Faculty of Science and Technology, Phetchabun Rajabhat University, Mueang, Phetchabun 67000, Thailand.
4Division of Physics, Faculty of Science and Technology, Phetchabun Rajabhat University, Mueang, Phetchabun 67000, Thailand.
Corresponding Author E-mail: sompong_s@rmutt.ac.th
DOI : http://dx.doi.org/10.13005/ojc/350304
Article Received on : 27-02-2019
Article Accepted on : 07-05-2019
Article Published : 14 May 2019
KDML 105 is the famous fragrant rice of Thailand. 2AP acts as the characteristic compounds in fragrant-rice cultivars. 2AP accumulation has been reported in relationship with proline content and the growth rate of rice. Several factors have been reported to affect the poline and 2AP contents. We determined the combinatorial effect of gamma irradiation and salt concentration on the 2AP and proline contents of 5 day-old KDML 105 rice-seedling. In the growth condition under 20 mM of NaCl concentration the proline content of gamma irradiated rice increased, compared to the gamma-irradiated rice treated without 20 mM of NaCl concentration. While, the shoot-length of 5 day-old gamma irradiated rice-seedlings under 20mM NaCl concentration cannot be observed when compared to gamma irradiated rice-seedling without salt concentration. The 2AP content of gamma irradiated rice-seedlings under 20 mM of NaCl concentration had approximately 2.6 to 3.1 times higher than in the growth condition without salt concentration of gamma irradiated rice-seedlings. Our results indicate that the combination of gamma irradiation technique and salt concentration can be used for improving the 2AP content in rice.
KEYWORDS:2-Acetyl-1-Pyrroline; Gamma Irradiation; KDML 105; Proline Content; Salt Stress
Download this article as:| Copy the following to cite this article: Sansenya S, Hua Y, Chumanee S, Sricheewin C. The Combination Effect of Gamma Irradiation and Salt Concentration on 2-Acetyl-1-Pyrroline Content, Proline Content and Growth of Thai Fragrant Rice (KDML 105). Orient J Chem 2019;35(3). |
| Copy the following to cite this URL: Sansenya S, Hua Y, Chumanee S, Sricheewin C. The Combination Effect of Gamma Irradiation and Salt Concentration on 2-Acetyl-1-Pyrroline Content, Proline Content and Growth of Thai Fragrant Rice (KDML 105). Orient J Chem 2019;35(3). Available from: https://bit.ly/2JoKNIb |
Introduction
The relationship between 2AP and proline contents of rice have been reported by many researchers. In the biosynthesis of 2AP, proline acts as the first precursory molecules.1-3 The researchers have reported that the accumulation of proline in rice may affect the accumulation of 2AP. Suprasanna et al.,4 found that when proline in callus of Basmati rice increased, the 2AP content also increased. The 2AP content also increased along with increasing of proline content in the callus and rice seedling of KDML105 rice.5 Poonlaphdecha et al.,6 also reported the increase of 2AP accumulation in rice under salt treatment due to the increase of proline content. Moreover, the 2AP content was related to the proline content when rice was treated with the reducing of solar intensity.7 On the contrary, Sansenya et al.,8 reported the 2AP accumulation in rice grain were not correlated with proline accumulation in rice grain of Thai local rice.
In recent years, many researchers have reported the effect of salt concentration on the 2AP accumulation in rice. The increase of salinity may affect the increase of 2AP content in grain of aroma rice variety.9 Poonlaphdecha et al.,6 reported that under salt treatment the increasing of 2AP content was obtained in rice grain but the salt concentration did not affect the other related compounds in the biochemical 2AP synthesis pathway such as GABA.
Gamma irradiation is the technique useful for inducing mutation to biological compounds of plant10-15. In rice, gamma irradiation has affected rice growth depending on gamma dose. Low gamma doses stimulated the rice growth, whereas at high gamma doses the inhibition of rice growth was observed.14-16 Gamma irradiation also affects the biological compounds of rice. Hwang et al.,17 found that the gamma irradiation caused some micronutrient content such as tocopherol in rice mutant line higher than in the control rice. Sansenya et al.,14 reported that the gamma irradiation affected the related biological compounds in 2AP biosynthesis pathway. According to the results, the gamma irradiation caused decrease of the GABA content, while the increasing of 2AP content was observed in Thai upland rice.
In this study, we report the combined effect of salt concentration and gamma irradiation to the rice growth and the related biological compounds in 2AP biosynthesis pathway such as proline and 2AP of KDML 105 rice.
Experimental
Plant Material and Gamma Irradiation
Seed of KDML 105 was obtained from the rice field in Roi-Et province, Thailand. The rice sample was sterilized with 0.1% NaClO and then washed with deionized water. The sterilized rice seeds was then roasted in hot air to decreased the moisture content to <13%.
For gamma irradiation, 100 grams were packed with polyethylene bag and then irradiated with the gamma ray intensity of 0 (control; without gamma irradiation), 20, 40, 60, 80, 100, 150, 300, and 500 Gy. We used137 Cs as the gamma source with the dose rate of 3 Gy/min at room temperature. The gamma irradiation was provided by Gamma Irradiation Service and Nuclear Technology Research Center, Faculty of Science, Kasetsart University, Thailand.
Effect of Germination Time on Proline Content of Non-Gamma Irradiated Rice Seedling
Fifty grams of non-gamma irradiated rice seeds were soaked in 100 mL of deionized water for 24 h. The rice seeds germinated on the germinating paper moistened by deionized water. The germination experiment was incubated at ambient temperature with 0 (without germination time), 1, 2, 3, 4, and 5 days for germination time, and during seed germination the experiments were sprayed with deionized water every 12 h. The germination seeds were harvested at the end of germination time at 0 h, 24 h (1 day), 48 h (2 days), 72 h (3 days), 96 h (4 days) and 120 h (5 days). Then the samples were kept at -20°C until proline quantified.
Effect of Salt Concentration on Proline Content and Growth of Non-Gamma Irradiated Rice-Seedlings (5 Days Old)
Fifty grams of non-gamma irradiated rice-seed were soaked in 100mL-salt solution of 0 (without NaCl concentration), 20, 40 ,60, 80 and 100mM of NaCl concentration for 24 h. The germination experiments were provided at the germination time that produced highest proline content (5 days). During of duration germination, the germination experiments were sprayed with salt solution of the same concentration as a soaking solution for every 12 h until 5 days. Next, sample processing was the same as experiment above.
Effects of Gamma Irradiation and Salt Concentration on 2AP Content, Proline Content of Rice Seedling (5 Days Old)
For the experiment without salt condition, the non-gamma irradiated rice-seed and gamma irradiated rice-seed were soaked in100 mL of deionized water for 24 h. Then, the germination experiments were sprayed with deionized water for every 12 h until 5 days.
For the experiment with salt condition, the non-gamma irradiated rice seed and gamma irradiated rice-seed were soaked in 20 mM salt solution for 24 h. Then, the germination experiments were sprayed with 20 mM salt solution for every 12 h until 5 days.
The rice-seedlings (5 days old) in both experiments were harvested and kept at -20°C until 2AP and proline quantified.
Effects of Gamma Irradiation and Salt Concentration on Plant Growth of Rice-Seedlings (5 Days Old)
For effect of salt concentration on plant growth. One hundred of non-gamma irradiated rice-seeds were soaked in salt solution including 0 (without NaCl concentration), 20, 40 ,60, 80 and 100 mM for 24 h. The germination experiments were carried out at the ambient temperature and sprayed with salt at the same concentration as soaking solution for every 12 h until 5 days. The rice seedlings at 5 days old were harvested and the rice shoot-lengths were measured.
For the effect of gamma irradiation on plant growth, one hundred of non-gamma irradiated rice-seeds and gamma irradiated rice-seeds were soaked in deionized water for 24 h. During duration germination, the experiment were sprayed with deionized water every 12 h until 5 days. Then, the rice-seedlings (5 days old) were harvested and the rice-shoot lengths were measured.
For effect of gamma irradiation and salt concentration on plant growth, one hundred of non-gamma irradiated rice-seeds and gamma irradiated rice-seeds were soaked in salt solution (20 mM NaCl) for 24 h. During duration germination, germinated rice were sprayed with 20mM NaCl solution for 5 days. Finally, the rice-seedlings (5 days old) were harvested and the rice shoot-lengths were measured.
Proline Quantification
Proline content of germinated rice was determined by the method of Sansenya et al.8 Rice samples of germinated rice-seeds were homogenized with CryoMill (Retch, Germany) by using liquid nitrogen. For proline determination, 0.3 g. of homogenized rice samples were extracted with 3% sulfosalicylic acid and before proline reaction the extraction solution were filtered with Syringe Filter (0.45 mm). The filtered solution for proline reaction was mixed with 3 mL of ninhydrin solution and 2 mL of glacial acetic acid, then the mixed reaction solution were boiled for 20 min. After boiling, the reaction solution were extracted with 4 mL of toluene. Next, the extracted solution was spectrophotometric at 520 nm. The proline concentration was calculated with the standard curve of proline: y = 0.413x-0.014 (R2 = 0.999).
2AP Quantification
The 2AP content of rice samples were determined by the modified method of Sansenya et al.8The homogenized rice samples for 2AP quantification was provided in the same way as in experiment for proline quantification. An agilent GC-MS (Agilent 7890A GC-7000 Mass Triple Quad) was used for the analysis. A 4 μL portion of the extract was injected into a DB-Wax (60 m × 0.25 mm i.d. × 0.25 μm film thickness) fused silica capillary column (J&W Scientific, Folsom, CA). The injector was set at 240°C, and split mode was applied with a split ratio of 5:1. Helium gas was used as the carrier gas with a constant flow rate of 1.5 mL/ min. The column oven was operated via temperature program: column temperature was isothermally maintained at 40°C for 2 min, programmed at a rate of 10°C/min to 100°C, then at a rate of 5°C/min to 150°C, and then at a rate of 30°C/min to 250°C; the column temperature was then maintained isothermally at 250°C for 15 min. The mass spectrometer was used in the electron ionization mode. The ion source temperature was set at 230°C, and ionization energy was set at 70 eV. Multi reaction monitoring (MRM) mode was set up to analyze 2AP. The precursor ion m/z 111 and the product ion m/z 83 were selected for 2AP with collision energy of 5 v. The mass spectrometer detection dwell time was 30 ms. Under these conditions, the retention times of 2AP were found to be 18.92 min. Quantification of 2AP was performed by measuring the area of ions at m/z 83, and the amounts of 2AP were calculated from its calibration curve, y = 19.68x-7.56, (R2 = 0.997).
Statistic Analysis
All the samples were expressed as the means ± standard deviation (means ± SD). The statistical significance of differences among experimental groups was evaluated by one-way analysis of variance (ANOVA). When the ANOVA revealed significant between-group differences, a post hoc analysis was performed using Duncan’s multiple-range test comparisons. The significance level was set at P < 0.05.
Results and Discussion
Effect of Salt Concentration on Proline Content and Growth of 5 days old Rice-Seedling
Our results show that the proline accumulation of rice was determined during duration of rice germination. The significant increase of proline content was observed at 1 day of duration germination compared with non-germinated rice (0 day). The proline content still increased after 1 day to 5 days during duration of germination (Table 1). In duration of seed germination, the soluble protein is degraded to the free amino acid by proteolytic enzyme.18 The proline is one of amino acid that is related to plant growth and plant differentiation.19 Our results were also performed according to the previous report, the proline content of 5 days old rice-seedlings was 9.5 times compared to that of non-germinated rice seed (0 day).
Table 1: The increasing of proline content during duration of rice germination.
|
Germination time (days) |
Proline content (mg/g) |
|
0 |
3.03 ± 0.06f |
|
1 |
10.04 ± 0.44e |
|
2 |
11.85 ± 0.42d |
|
3 |
13.70 ± 0.29c |
|
4 |
20.89 ± 0.63b |
|
5 |
28.88 ± 0.19a |
± indicates the standard deviation from means (n=3).
The error bar indicates the standard deviation from means (n = 3). The same letter indicates no significant difference (Dancan, p>0.05).
Many researchers reported that the proline was one of the amino acids that respond to the environmental stress such as salt stress, dough stress and radiation stress.20-26 Our result shows the correlation of proline accumulation and plant growth of 5 days old rice-seedlings under salt condition. Rice-seedlings (5 days old) under salt condition reveal that the proline content increased at low salt-concentrations (20 and 40mM NaCl concentration) (Fig. 1). The highest proline content of 5 days old rice-seedlings was observed at 20 mM NaCl concentration and approximately 2 times compared to 5 days old rice seedling without salt concentration (0mM NaCl concentration). The significant decreasing of proline content was observed when rice was treated with high salt-concentration and approximately 3.4 times decreasing, compared with highest proline content of 5 days old rice-seedlings treated with 20mM salt concentration.
The correlation of proline content and rice-shoot-lengths of 5 days old rice-seedlings was found in all salt conditions. The increasing of rice shoot-lengths was observed when the proline content increased. While, the low concentration of proline content was obtained from shorter rice shoot-lengths. Moreover, the results show that we may obtain highest proline content and highest rice shoot-lengths from low salt-concentrations (20 mM salt concentration). The rice shoot-lengths and proline content were decreased after rice was treated with high salt-concentrations (Fig. 1 and 2). This results agree with Kibria,27 the proline content significantly increased in BRRI dhan28 rice cultivar under low salt concentration (20 and 40 mM salt concentration), while at above 60mM salt concentration it caused decrease in the proline content of rice. For other rice cultivars, the high salt concentration caused increasing the proline content of rice such as BRRI dhan 47, Binadhan-8 and Binadhan-10.27 Our result and other report indicate that in different rice cultivars, the level of salt tolerance might be different and the maintenance of proline content should also be different.
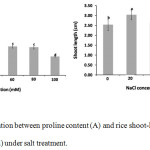 |
Figure 1: The correlation between proline content (A) and rice shoot-length (B) of rice seedlings (5 days old) under salt treatment. |
The error bar indicates the standard deviation of means (n = 3 for proline content and n = 6 for rice shoot-length ). The same letter indicates no significant difference (Dancan, p>0.05).
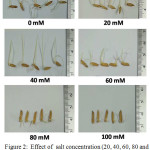 |
Figure 2: Effect of salt concentration (20, 40, 60, 80 and 100mM NaCl) on rice shoot-lengths of rice-seedlings (5 days old). |
The 5 days old rice seedlings show increase of rice shoot-length under 20mM NaCl concentration compared to rice shoot-lengths of control rice and 40 to 100 mM NaCl concentration.
Effect of Salt Concentration on the Proline Content and Growth of 5 days old Gamma Irradiated Rice-Seedlings
Gamma ray has an effect on living cells of organism depending on the gamma dose.28 The gamma ray has generated free radical and this induced mutation in plant cells.29-32 We determined the effect of gamma irradiation on the proline content and the plant growth of 5 days old rice seedlings with and without salt concentration (20mM NaCl). Proline content of gamma irradiated rice-seedlings with and without 20 mM NaCl increased after treating with 40 and 60 Gy with respect to 0 Gy. At high gamma doses (80 to 500 Gy) the proline content decreased continuously in both conditions (with and without 20mM NaCl concentration) (Fig. 3A). The largest decrease of proline content of gamma irradiated rice seedlings were observed in case of without 20mM NaCl, whereas the slight decrease of proline content was observed in gamma irradiated rice-seedlings treated with 20 mM NaCl. The highest and the lowest proline contents of gamma irradiated rice-seedlingsin both conditions were obtained from 20 Gy for highest proline content and 500 Gy for lowest proline content respectively. The trend of proline content of gamma irradiated rice-seedlings with 20mM NaCl had higher proline content of gamma irradiated rice-seedlings than in case of without 20mM NaCl of all gamma dose. Gamma ray also affects the shoot length of 5 days old rice seedlings. Compared with the control, the gamma irradiated rice-seedlings treated without 20mM NaCl concentration at 20 Gy to 150 Gy showed significant differences in their shoot lengths. The highest shoot lengths were observed at 20Gy gamma dose (Fig. 3 B; gay bar and Fig. 4). While, their shoot lengths were lower in the 300 Gy and 500 Gy gamma irradiated rice-seedlings than in the control rice (Fig. 3 B; gray bar and Fig. 4). Interestingly, for the gamma irradiated rice-seeds treated with 20mM NaCl concentration, the germination can be observed in duration of 3 days, but the transformation from germinated rice-seeds to rice-seedlings cannot be observed in case of gamma irradiation at 20 Gy to 500 Gy with respect to the control set (Fig. 3 B; dark gay bar and Fig. 5). The stimulation via low gamma-irradiation dose under salt condition to increased roline content of 5 days old rice seedlings of our results are similar to the study of El-Beltagi et al.,24 who found that all salt concentrations under 50Gy gamma dose induced the proline content in the shoots of Cowpea plants. Many reports show that salt condition affects the proline level in plants cell.22,33,34 In general, proline has many behaviors during salt stress such as accumulation of energy reservoir, nitrogen source, water storage, pH stability in the cell and hydroxyl radical scavenging.19,22,26,35-37 Gamma rays may also induce the proline content under low gamma dose described by Al-Enezi and Al-Khayri.38 Gamma irradiation produces the radical having many effects on plant cells. The increase of proline content of gamma irradiated rice-seedlings under salt condition compared to the gamma irradiated rice seedlings treated without salt concentration may affect plant growth under combination stress (Fig. 3A). At low gamma doses (20 Gy to 150 Gy) the shoot lengths of 5 days old rice-seedlings are higher than those of control rice (Fig. 3B gray bar). However, high gamma doses (300Gy to 500 Gy) seem to inhibit the plant growth of 5 days old rice seedlings (Fig. 3B; gray bar and Fig. 4). Many researchers have been reported the stimulation of plant growth by low doses of gamma-irradiation. Chandrashekar et al.,23 reported the higher shoot lengths of Terminalia arjuna Roxb than the control set under low doses of gamma-irradiation. Sansenya et al.,15 also reported the stimulation of low gamma irradiation doses on shoot lengths of Thai black glutinous rice. Moreover, under combination stress (salt concentration and gamma irradiation) it inhibited plant growth of 5 days old rice seedling of all gamma doses with respect to the control rice (Fig. 3B dark gray bar and Fig. 5). Previous researchers also reported the inhibition of growth of gamma irradiated plants under salt condition. The shoot lengths of gamma irradiated Tarrom Hashemi (Oryza sativa L.) had decreased under salt condition. More, the decreasing of germination percentage were also observed in gamma irradiated rice-seeds under salt condition (Tarrom Hashemi)21. The inhibition of growth of gamma irradiated plants under salt condition may cause many damages in the genome of plant.39 Moreover, while plant growth under salt condition had effect on the loss of water content from plant and inducing toxicity in the plant cell may cause inhibition of the growth rate of plant.40
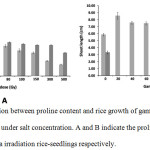 |
Figure 3: The correlation between proline content and rice growth of gamma irradiation rice-seedling (5 days old) under salt concentration. A and B indicate the proline content and rice shoot-length of gamma irradiation rice-seedlings respectively. |
The gray bar indicates gamma irradiation rice-seedlings treated without salt concentration, while the dark gray bar indicates gamma irradiation rice-seedlings treated with 20mM salt concentration. The error bar indicates the standard deviation of means (n = 3 for proline concentration and n = 6 for rice shoot-length).
 |
Figure 4: Effect of gamma irradiation (20, 40, 60, 80, 100, 150, 300 and 500 Gy) on rice shoot-lengths of rice-seedlings (5 days old). 0 Gy means treating with no gamma irradiation. |
 |
Figure 5: The combination effect of gamma irradiation (20, 40, 60, 80, 100, 150, 300 and 500 Gy) and salt concentration (20mM NaCl) on rice shoot-lengths of rice-seedlings (5 days old). 0 Gy mean treating with no gamma irradiation. |
Effect of Salt Concentration on 2AP Content of 5 days old Gamma Irradiated Rice Seedlings
2AP is the aroma compounds that acts as a characteristic compound for fragrant rice.2,8,41,42 This compound is synthesized via L-proline metabolism and is associated with inactive betiane aldehyde dehydrogenase (Badh2)2. The level of 2AP accumulation in rice may be caused by several environmental factors such as salt stress, dough stress, reducing of solar intensity and gamma irradiation.6,7,14,15,43 Our results show that the gamma irradiation affects increasing of 2AP content of 5 days old rice seedlings of all gamma dose compared with control rice (Fig. 6 gray bar). The 2AP content of gamma-irradiated rice increases when gamma dose increases (20 Gy to 300 Gy), whereas at 500 Gy the 2AP content of gamma irradiated rice decreased with respect to other gamma doses (20 Gy to 300 Gy) (Fig. 6 gray bar). We have determined the effect of salt concentration on 2AP content of 5 days old gamma irradiated rice-seedling. The 2AP content of non-gamma-irradiated rice (0 Gy) that grew under 20mM NaCl concentration had 6.7-fold higher than in case of non-gamma-irradiated rice (0 Gy) treated without 20mM NaCl concentration (Fig. 6). While other 5 days old rice seedlings treated with 20Gy to 500Gy with 20mM NaCl concentration also had 2AP content approximately 2.6 to 3.1-times higher than in case of gamma irradiated rice seedling without 20mM NaCl concentration. Moreover, the 2AP content of 5 days old rice seedlings treated with gamma irradiation and with 20mM NaCl concentration gradually increased at 20 Gy to 80 Gy, whereas the 2AP content continuously decreased at 100 Gy to 500 Gy compared to control rice (Fig. 6 dark gray bar). Salt and gamma irradiation condition have been regarded as factors that affect the 2AP accumulation in rice. The increase of 2AP content under salt concentration has been reported by Poonlaphdecha et al.,6 and Yoshihashi et al.43 The gamma irradiation also affected the increasing of the 2AP content in germinated rice and gamma irradiated rice-seeds14,15. Proline is the precursor for 2AP synthesis metabolism, under salt condition it has been reported to stimulate the proline content in rice. Increases of proline content in gamma irradiated rice were also reported.21 Fig. 3A of our result also shows that the proline content of gamma irradiated rice-seedlings with 20mM salt concentration condition has increased with respect to controlled rice. The increase of proline content under combination factors might explain the increasing of 2AP content of gamma-irradiated rice under 20mM NaCl concentration.
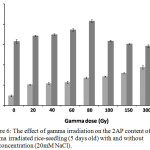 |
Figure 6: The effect of gamma irradiation on the 2AP content of gamma irradiated rice-seedling (5 days old) with and without salt concentration (20mM NaCl). |
The gray bar indicates gamma irradiated rice-seedlings treated without salt concentration, while the dark gray bar indicates gamma irradiated rice-seedlings treated with 20mM salt concentration. The error bar indicates the standard deviation from means (n = 3).
Conclusion
Poline content of KDML 105 rice gradually increased from 1 to 5 days compared to non-germinated rice during germination. The proline content and the shoot-length of 5 days old rice-seedlings had highest value for 20mM NaCl concentration, whereas high salt concentration seemed to inhibit proline content and rice shoot-length. The proline content of 5 days old gamma irradiated rice-seedlings under 20mM NaCl concentration had higher value than that of gamma-irradiated rice treated without salt concentration. Salt concentration has inhibited the rice shoot-lengths of gamma-irradiated rice compared to the gamma-irradiated rice growth in the condition of without salt concentration. In growth condition with 20mM NaCl concentration, the gamma irradiated rice-seedling had the 2AP content higher than in case of no salt concentration.
Acknowledgements
This research was supported by the National Research Council of Thailand (Grant 2560A16502109) and Rajamangala University of Technology Thanyaburi. We are also grateful to Kadsada Sala for providing plant samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Buttery, R.G.; Ling, L.C.; Juliano, B.O.; Turnbaugh, J.G. Cooked rice aroma and 2-acetyl-1-pyrroline. J Agric Food Chem. 1983, 318, 23-826.
- Bradbury, L.M.; Fitzgerald, T. L.; Henry, R. J., Jin, Q., & Waters, D. L. The gene for fragrance in rice. Plant Biotechnol J. 2005, 3, 363-370.
- Wakte, K.; Zanan, R.; Hinge, V.; Khandagale, K.; Nadaf, A.; Henry, R. Thirty‐three years of 2‐acetyl‐1‐pyrroline, a principal basmati aroma compound in scented rice (Oryza sativa L.): a status review. J. Sci. Food Agric. 2017, 97, 384-395.
- Suprasanna, P.; Ganapathi, T.R.; Ramaswamy, N.K.; Surendranathan, K.K.; Rao, P.S. Aroma synthesis in cell and callus cultures of rice. Rice Genet News. 1998, 15, 123-125.
- Yoshihashi , T.; Huong, N.T.; Inatomi, H. Precursors of 2-acetyl-1-pyrroline, a potent flavor compound of an aromatic rice variety. J Agric Food Chem. 2002, 50, 2001- 2004.
- Poonlaphdecha, J.; Maraval, I.; Roques, S.; Audebert, A.; Boulanger, R.; Bry, X.; Gunata, Z. Effect of timing and duration of salt treatment during growth of a fragrant rice variety on yield and 2-Acetyl-1-pyrroline, proline, and GABA levels. J Agric Food Chem. 2012, 60, 3824-3830.
- Mo, Z.; Li, W.; Pan, S.; Fitzgerald, T.L.; Xiao, F.; Tang, Y.; Wang, Y.; Duan, M.; Tian, H.; Tang, X. Shading during the grain filling period increases 2-acetyl-1-pyrroline content in fragrant rice. Rice. 2015, 8, 9.
- Sansenya, S.; Hua, Y.; Chumanee, S. The Correlation between 2-Acetyl-1-pyrroline Content, Biological Compounds and Molecular Characterization to the Aroma Intensities of Thai Local Rice. J Oleo Sci. 2018, 67, 893-904.
- Gay, F.; Maraval, I.; Roques, S.; Gunata, Z.; Boulanger, R.; Audebert, A.; Mestres, C. Effect of salinity on yield and 2-acetyl-1-pyrroline content in the grains of three fragrant rice cultivars (Oryza sativa L.) in Camargue (France). Field Crops Res. 2010, 117, 154-160.
- Killion, D.D.; Constantin, M. J.; Siemer, E. G. Acute gamma irradiation of the soybean plant: effects of exposure, exposure rate and developmental stage on growth and yield. Radiat Bot. 1971, 11, 225-232.
- Singh, B.; Datta, P.S. Gamma irradiation to improve plant vigour, grain development, and yield attributes of wheat. Radiat Phys Chem. 2010, 79, 139-143.
- Marcu, D.; Damian, G.; Cosma, C.; Cristea, V. Gamma radiation effects on seed germination, growth and pigment content, and ESR study of induced free radicals in maize (Zea mays). J Biol Phys. 2013, 39, 625-634.
- Minisi, F.A.; El-mahrouk, M.E.; Rida, M.E.F.; Nasr, M.N. Effects of gamma radiation on germination, growth characteristics and morphological variations of Moluccella laevis L. Am.-Eurasian J. Agric. Environ. Sci. 2013, 13, 696-704
- Sansenya, S.; Hua, Y.; Chumanee, S.; Phasai, K.; Sricheewin, C. Effect of Gamma Irradiation on 2-Acetyl-1-pyrroline Content, GABA Content and Volatile Compounds of Germinated Rice (Thai Upland Rice). Plants, 2017, 6, 18
- Sansenya, S.; Hua, Y.; Chumanee, S.; Winyakul, C. Effect of gamma irradiation on the 2-acetyl-1-pyrroline content during growth of Thai black glutinous rice (Upland rice). Aust J Crop Sci. 2017, 11, 631.
- Sasikala, R.; Kalaiyarasi, R. Sensitivity of rice varieties to gamma irradiation. Electron. j. plant breed. 2010, 1, 885-889.
- Hwang, J.E.; Ahn, J.W.; Kwon, S.J.; Kim, J.B.; Kim, S.H.; Kang, S.Y.; Kim, D.S. Selection and molecular characterization of a high tocopherol accumulation rice mutant line induced by gamma irradiation. Mol Biol Rep. 2014, 41, 7671-7681.
- Kirmizi, S.; Güleryüz, G. Protein mobilization and proteolytic enzyme activities during seed germination of broad bean (Vicia faba L.). Z. Naturforsch. C Bio. Sci. 2006, 61, 222-226.
- Kishor, K.; Polavarapu, B.; Hima Kumari, P.; Sunita, M.S.L.; Sreenivasulu, N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front Plant Sci. 2015, 6, 544.
- Mattioli, R.; Costantino, P.; Trovato, M. Proline accumulation in plants: not only stress. Plant Signal Behav. 2009, 4, 1016-1018,
- Dehpour, J.A.A.; Gholampour, M.; Rahdary, P.; Jafari, T.M.R.; Hamdi, S.M.M. Effect of gamma irradiation and salt stress on germination, callus, protein and proline in rice (Oryza sativa L.), Iran. J Plant Physiol. 2011, 1, 251‐256.
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: a review. Plant Signal Behav. 2012, 7, 1456-1466.
- Chandrashekar, K.R.; Somashekarappa, H.M.; Souframanien, J. Effect of gamma irradiation on germination, growth, and biochemical parameters of Terminalia arjuna Roxb. Radiat Prot Environ. 2013, 36, 38
- El-Beltagi, H.S.; Mohamed, H. I.; Mohammed, A.H.M.; Mogazy, A.M. Physiological and Biochemical Effects of γ-Irradiation on Cowpea Plants (Vigna sinensis) under Salt Stress. Not Bot Horti Agrobot Cluj Napoca. 2013, 41, 104-114.
- Jaarsma, R.; de Vries, R.S.; de Boer, A.H. Effect of salt stress on growth, Na+ accumulation and proline metabolism in potato (Solanum tuberosum) cultivars. PLoS One. 2013, 8, e60183
- Ali, H.; Ghori, Z.; Sheikh, S.; Gul, A. Effects of gamma radiation on crop production. In Crop Production and Global Environmental. Springer, Cham. 2015, pp. 27-78.
- Kibria, M.G. Physiological and biochemical responses of salt-sensitive and salt-tolerant rice genotypes to salt stress. Mymensingh, Bangladesh: Bangladesh Agricultural University, MSc thesis. 2015.
- Kim, J.H.; Chung, B.Y.; Kim, J.S.; Wi, S.G. Effects ofin Planta gamma-irradiation on growth, photosynthesis, and antioxidative capacity of red pepper (Capsicum annuum L.) plants. J Plant Biol. 2005, 48, 47-56.
- Gunkel, J.E.; Sparrow, A.H. Ionizing radiations: Bio‑chemical, physiological and morphological aspects of their effects on plants. Encyc Plant Phys. 1961, 16, 555‑611.
- Kovacs, E.; A. Keresztes, Effect of gamma and UV‑B/C radiation on plant cell. Micron. 2002, 33, 199‑210.
- Kim, J.H.; Baek, M.H.; Chung, B.Y.; Wi, S.G.; Kim, J.S. Alterations in the photosynthetic pigments and antioxidant machineries of red pepper (Capsicum annuum L.) seedlings from gamma‑irradiated seeds. J Plant Biol. 2004, 47, 314‑21.
- Wi, S.G.; Chung, B.Y.; Kim, J.S.; Kim, J.H.; Baek, M.H.; Lee, J.W.; Kim, Y.S. Effects of gamma irradiation on morphological changes and biological responses in plants. Micron. 2007, 38, 553-564.
- Benhassaini, H.; Fetati, A.; Hocine, A.K.; Belkhodja, M. Effect of salt stress on growth and accumulation of proline and soluble sugars on plantlets of Pistacia atlantica Desf. subsp. atlantica used as root stocks. Biotechnol. agron. soc. environ. 2012, 16, 159.
- Gharsallah, C.H.; Fakhfakh, Grubb, D.; Gorsane, F. Effect of salt stress on ion concentration, proline content, antioxidant enzyme activities and gene expression in tomato cultivars. AoB Plants. 2016, 27, 8.
- Irigoyen, J.J.; Einerich, D.W.; Sánchez‐Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd). Plants. Physiol. Plant. 1992, 84, 55-60.
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: an overview. Arch Biochem Biophys. 2005, 444, 139-158.
- Kuznetsov, V.V.; Shevyakova, N.I. Polyamines and stress tolerance of plants. Plant Stress. 2007, 1, 50-71.
- Al-Enezi, N.A.; Al-Khayri, J.M. Effect of X-irradiation on proline accumulation. Growth and water content of date palm (Phoenix dactylifera L.) seedlings. J Biol Sci. 2012, 12, 146-153.
- Preuss, S.B.; Britt, A.B. A DNA-damage-induced cell cycle checkpoint in Arabidopsis. Genetics. 2003, 164, 323-334.
- Ghoulam, C.; Fares, K. Effect of salinity on seed germination and early seedling growth of sugar beet (Beta vulgaris L.). Seed Sci. Technol. 2001, 29, 357-364,
- Buttery, R.G.; Turnbaugh, J.G.; Ling, L.C. Contribution of volatiles to rice aroma. J Agric Food Chem. 1988, 36, 1006-1009.
- Huang, T.C.; Teng, C.S.; Chang, J.L.; Chuang, H.S.; Ho, C.T.; Wu, M.L. Biosynthetic mechanism of 2-acetyl-1-pyrroline and its relationship with Δ1- pyrroline-5-carboxylic acid and methylglyoxal in aromatic rice (Oryza sativa L.) callus. J Agric Food Chem. 2008, 56, 7399-7404.
- Yoshihashi, T.; Nguyen, T.T.H.; Kabaki, N. Area dependency of 2-acetyl-1-pyrroline content in an aromatic rice variety, Khao Dawk Mali 105. Jpn Agric Res Q. 2004, 38, 105-109.

This work is licensed under a Creative Commons Attribution 4.0 International License.
![]()
A New Edition of Web of Science
Journal Impact Factor
2022: 0.5
Five Year: 0.8
Journal is Indexed in

Cabells Whitelist
![]()


