Phytochemical Profiling of Medicinal Plant Grown Under Different Soil Conditions
 , Harvinder Kaur Sidhu2, Nileema Pandey3, Ashok Kumar Koshariya4*, Shivaraj Kumar Verma5, Indra Jeet6, Sivaraman Ramanarayanan7 and Diksha Sinha8
, Harvinder Kaur Sidhu2, Nileema Pandey3, Ashok Kumar Koshariya4*, Shivaraj Kumar Verma5, Indra Jeet6, Sivaraman Ramanarayanan7 and Diksha Sinha81Department Agricultural Engineering, Krishi Vigyan Kendra, Puri, OUAT, Odisha, India.
2Agriculture and Life Sciences, Desh Bhagat University, Mandi Gobindgarh, Punjab, India.
3Floriculture and Landscape, MGUVV, Sakara, Patan, Durg, Chhattisgarh, India.
4Plant Pathology, Gurughasidas Regional Agriculture Research Station, IGKV, Kawardha, Chhattisgarh, India.
5Department of Horticulture, Faculty of Agriculture, Udai Pratap College, Varanasi (U.P. ), India.
6ICAR-RCER, Krishi Vigyan Kendra, Ramgarh, Jharkhand, India.
7Department of Veterinary Pharmacology and Toxicology, College of Veterinary and Animal Sciences, Kishanganj, Bihar Animal Sciences University, Bihar, India.
8MoFECC, Government of Jharkhand, India.
Corresponding Author E-mail:ashok.koshariya@gmail.com
DOI : http://dx.doi.org//10.13005/ojc/420327
Download this article as:
![]()
The therapeutic efficacy of medicinal plants depends critically on the consistency of their phytochemical profiles—the concentrations and ratios of alkaloids, flavonoids, terpenes, saponins, and phenolic acids. However, these secondary metabolites are not fixed traits but dynamic outputs of the plant’s adaptive responses to edaphic (soil) conditions. This review provides a comprehensive mechanistic framework linking soil parameters to phytochemical biosynthesis, emphasizing that soil functions as a “chemical reactor” where physical (texture, structure, water-holding capacity), chemical (pH, electrical conductivity, redox potential), biological (microbiome composition, mycorrhizal associations), and nutrient (N, P, K, S, Fe, Zn, Mn, Cu) parameters collectively determine metabolic fate. Nine mechanistic pathways are delineated: general stress-induced defense responses (jasmonate-mediated), nitrogen-driven trade-offs between N-containing alkaloids and C-based phenolics, phosphorus-limitation upregulation of flavonoids and organic acids, potassium modulation of terpenoid/saponin biosynthesis, soil pH as a master variable controlling nutrient solubility and ion antagonism, salinity-induced accumulation of osmoprotectants (proline, glycine betaine) and phenolic acids, heavy metal stress triggering chelator (phytochelatin, glucosinolate, flavonoid) synthesis, water stress (drought/flooding) mediating abscisic acid regulation of terpenes and lignans, and soil microbiome signaling through microbial-associated molecular patterns and volatile organic compounds. Five case studies illustrate species-specific responses: Panax (ginseng) ginsenoside profiles governed by pH and microbiome; Artemisia annua artemisinin modulated by nitrogen form and controlled deficit irrigation; Cannabis sativa cannabinoid/terpene plasticity under salinity and metal stress; Glycyrrhiza (licorice) glycyrrhizin affected by soil texture and sulfate-dominated salinity; and Hypericumperforatum hypericin/pseudohypericin determined by light–nutrient interactions. Translational applications include precision agriculture for standardized herbal products, conservation of chemotypes under habitat change, soil-stress elicitation for novel metabolite discovery, and integration of edaphic metadata into pharmacopoeial standards. Persistent challenges remain—nonlinear interactions, cost of routine soil analysis, post-harvest processing overriding edaphic effects, regulatory reliance on marker compounds rather than soil-based certification, and economic trade-offs between phytochemical content and biomass yield. Future directions emphasize high-throughput sensor networks, synthetic microbial consortia, predictive modeling under climate change, and mandatory soil metadata reporting. Bridging soil science with phytochemical analytics transforms quality control from reactive testing to predictive, soil-informed design—essential for delivering consistent, safe, and effective herbal therapeutics.
KEYWORDS:Artemisinin; Cannabinoids; Edaphic Factors; Ginsenosides; Glycyrrhizin; Hypericin; Medicinal Plants; Phytochemical profiling; Rhizosphere; Soil Parameters; Secondary Metabolites;
Introduction
The Significance of Phytochemical Consistency in Medicinal Plants
The therapeutic efficacy of medicinal plants hinges not merely on the presence of bioactive compounds but on their reproducible, predictable concentrations across batches, harvests, and geographic locations. Phytochemical consistency—the maintenance of stable qualitative and quantitative profiles of secondary metabolites such as alkaloids, flavonoids, terpenes, and phenolic acids—represents the cornerstone of herbal medicine safety, efficacy, and regulatory compliance. Unlike synthetic pharmaceuticals, which are manufactured under tightly controlled conditions to ensure batch-to-batch uniformity, medicinal plants are biological systems inherently susceptible to environmental fluctuation. This variability poses profound clinical risks: a Digitalis purpurea preparation with marginally elevated cardiac glycosides can induce fatal arrhythmias, while Artemisia annua with suboptimal artemisinin content may fail to clear malarial parasites, promoting drug resistance. Beyond clinical outcomes, inconsistent phytochemical profiles undermine reproducibility in pharmacological research, confound mechanistic studies, and erode consumer trust in herbal therapeutics. Regulatory frameworks such as the European Medicines Agency’s guidelines on herbal medicinal products and the World Health Organization’s monographs increasingly demand validated analytical methods to quantify marker compounds and establish phytochemical fingerprints. However, the deeper challenge lies in understanding why inconsistency arises in the first place. Phytochemical profiles are not random; they are the plant’s adaptive responses to specific ecological pressures, including light intensity, temperature, water availability, herbivory, and crucially, soil conditions. Without controlling or accounting for these drivers, even rigorous quality control becomes reactive rather than predictive. Thus, achieving true consistency requires moving beyond post-harvest standardization to a systems-level understanding of how edaphic and environmental factors shape metabolic fate. This recognition sets the stage for investigating soil not as an inert growth medium but as a dynamic determinant of plant chemistry.
 |
Figure 1: Root System and Rhizosphere Interactions Click here to View Figure |
Soil as a Determinant of Plant Growth and Metabolic Fate
Soil functions as far more than a physical anchor for root systems; it is a complex, living matrix of minerals, organic matter, water, gases, and a diverse microbiome that collectively orchestrates plant development and secondary metabolism. The edaphic environment directly influences nutrient bioavailability, water uptake, root architecture, and stress signaling—all of which modulate biosynthetic pathways responsible for phytochemical production. Macronutrients such as nitrogen, phosphorus, and potassium play foundational roles: nitrogen availability, for instance, governs the allocation between primary metabolism (growth) and secondary metabolism (defense). High nitrogen typically suppresses carbon-based secondary metabolites like phenolics and terpenoids while favoring nitrogen-containing alkaloids, though the exact response varies by species and compound class. Phosphorus deficiency, conversely, often elevates flavonoid and anthocyanin accumulation as part of oxidative stress mitigation. Beyond macronutrients, trace elements including iron, zinc, manganese, and boron serve as cofactors for enzymes central to terpene biosynthesis, lignin formation, and antioxidant defense. Soil pH exerts overarching control by determining the solubility and mobility of these elements; acidic soils may mobilize toxic aluminum while rendering molybdenum unavailable, each perturbation triggering distinct shifts in phytochemical profiles. Perhaps most critically, the rhizospheremicrobiome—bacteria, fungi, protozoa, and viruses living in intimate association with roots—acts as an extended metabolic genome. Arbuscularmycorrhizal fungi enhance phosphorus uptake and frequently upregulateterpenoid and phenolic biosynthesis. Endophytic bacteria can produce their own bioactive metabolites or elicit plant defense pathways through volatile signaling. Soil texture and structure influence water-holding capacity and aeration, thereby modulating drought stress responses. A clay-rich soil that becomes waterlogged will induce hypoxia, prompting accumulation of stress-related alkaloids and polyamines, whereas a sandy soil prone to rapid drainage may trigger osmotic adjustment via proline and soluble phenolics. Importantly, these soil factors do not operate in isolation; they interact through complex networks of synergistic, antagonistic, or threshold effects. Consequently, two plants of the same species growing just meters apart on different soil patches can exhibit divergent phytochemical fingerprints. This edaphic-driven variability is neither noise nor nuisance—it is a fundamental biological reality that any serious effort toward phytochemical consistency must confront directly.
Bridging Mechanisms and Analytical Strategies
Recognizing that soil governs phytochemical profiles through mechanistically distinct yet interconnected pathways, the central objective of this work is to bridge the conceptual and methodological gap between pedological drivers and analytical quality control. Traditional approaches have largely operated in disciplinary silos: soil scientists characterize nutrient cycles and microbial communities without linking these parameters to specific compound classes, while analytical chemists develop increasingly sophisticated HPLC-MS and NMR methods to quantify marker compounds without systematically correlating results to edaphic variables. This fragmentation produces descriptive rather than predictive knowledge—we can measure inconsistency but cannot reliably anticipate or prevent it. The present scope therefore integrates three complementary domains: (1) mechanistic elucidation of how specific soil parameters modulate particular biosynthetic pathways, (2) development of targeted and untargeted analytical strategies capable of capturing edaphic influences on phytochemical profiles, and (3) translation of these insights into practical frameworks for cultivation, harvesting, and quality assurance. Mechanistically, we prioritize several key pathways: the shikimic acid pathway yielding phenolic acids and flavonoids, the mevalonate and methylerythritol phosphate pathways producing terpenoids, and the alkaloid biosynthetic routes derived from amino acid precursors. For each, we examine how soil nitrogen, phosphorus, potassium, pH, organic matter, and microbiome composition alter enzyme activities, gene expression, and substrate availability. Analytically, we advocate for hybrid strategies that combine high-resolution metabolomics for discovery-phase fingerprinting with targeted quantitative assays for validated marker compounds. Chemometric tools—principal component analysis, partial least squares discriminant analysis, and machine learning algorithms—are essential for modeling relationships between soil edaphic parameters (predictor variables) and phytochemical profiles (response variables). Such models can identify which soil factors exert the strongest influence on therapeutic consistency, enabling site-specific cultivation recommendations or soil amendment protocols. Furthermore, we explore emerging technologies including portable near-infrared spectroscopy for real-time field assessment of phytochemical status and DNA metabarcoding for rapid characterization of rhizosphere communities. The ultimate objective is a predictive framework wherein knowledge of a soil’s physical, chemical, and biological properties allows accurate forecasting of a medicinal plant’s phytochemical profile prior to harvest. This shift from reactive testing to proactive design represents the frontier of phytochemical consistency. By systematically bridging the mechanisms by which soil determines metabolic fate with the analytical strategies required to measure and manage this relationship, we aim to provide both theoretical insights and actionable tools for researchers, growers, and quality control laboratories. The following chapters will elaborate each component in detail, moving from foundational principles to case studies and finally to integrated protocols for edaphically-informed phytochemical quality assurance.
The Rhizosphere as a Chemical Reactor: Key Soil Parameters
The rhizosphere—the narrow zone of soil directly influenced by root exudates and associated microorganisms—functions as a dynamic chemical reactor where physical, chemical, and biological parameters converge to govern nutrient bioavailability, stress signaling, and ultimately the phytochemical output of medicinal plants. Understanding this reactor’s operating conditions is essential because even subtle variations in soil properties can cascade into dramatic shifts in secondary metabolite profiles. Unlike bulk soil, the rhizosphere is characterized by steep gradients in pH, oxygen concentration, organic carbon, and microbial activity over millimeter to sub-millimeter scales, creating microhabitats that differentially regulate plant metabolic responses. Four categories of soil parameters—physical, chemical, biological, and nutrient dynamics—operate interdependently within this reactor, and their integrated effects determine whether a medicinal plant produces consistent, therapeutically relevant concentrations of bioactive compounds or exhibits problematic batch-to-batch variability.
Physical Properties: Texture, Structure, and Water-Holding Capacity
Soil texture—the relative proportions of sand, silt, and clay particles—establishes the fundamental physical framework within which the rhizosphere reactor operates. Sandy soils, dominated by large particles, exhibit rapid drainage and low surface area for nutrient retention, forcing roots to expend energy on water and nutrient foraging rather than secondary metabolism; this often results in elevated concentrations of stress-induced phenolics and proline but reduced biomass and variable alkaloid profiles. Clay-rich soils, conversely, provide extensive surface area for cation exchange and water retention but may become hypoxic when saturated, triggering anaerobic metabolism and accumulation of stress-related polyamines and volatile aldehydes that can alter essential oil composition in aromatic medicinal plants like Salvia miltiorrhiza or Lavandulaangustifolia. Soil structure—the aggregation of primary particles into peds—modifies these textural effects by creating pore networks that govern root penetration, gas exchange, and water flow. Well-structured, granular soils with balanced macro- and micro-porosity promote uniform root distribution and stable rhizosphere conditions, favoring consistent phytochemical profiles. Compacted or structureless soils restrict root exploration, concentrate exudates in localized zones, and promote pathogenic microbial proliferation, all of which induce erratic secondary metabolite accumulation. Water-holding capacity, a direct consequence of texture and organic matter content, acts as a master variable: cyclic drought and rewetting events trigger jasmonic acid signaling cascades that upregulateterpenoid and alkaloid biosynthetic genes, but excessive soil moisture suppresses these responses while promoting root rot and associated mycotoxin contamination. For medicinal plants such as Panax ginseng, which requires well-drained yet consistently moist conditions, deviations in water-holding capacity of even 10-15% can shift ginsenoside profiles from predominantly protopanaxadiol to protopanaxatriol forms, with direct implications for neuroprotective versus anti-inflammatory activities. Thus, physical properties do not operate passively; they set the boundary conditions for the chemical and biological processes that directly mediate phytochemical biosynthesis.
 |
Figure 2: Soil Parameters and Plant Signaling Pathways Click here to View Figure |
Chemical Properties: pH, Electrical Conductivity, and Redox Potential
Within the rhizosphere reactor, soil pH functions as the primary chemical governor, determining the solubility, mobility, and plant availability of virtually every nutrient and metal ion. In acidic soils (pH 4.0–5.5), aluminum and manganese become soluble to phytotoxic levels, while phosphorus is sequestered by iron and aluminum oxides, and molybdenum availability plummets. Medicinal plants respond to aluminum toxicity by exuding citrate and malate from roots—organic acids that chelate the toxic ion but also drain carbon skeletons away from phenylpropanoid and terpenoid pathways. Simultaneously, low pH often increases the concentration of protonated alkaloids within root tissues, altering the alkaloid profile in species like Catharanthusroseus (vinca alkaloids) or Colchicum autumnale (colchicine). In alkaline soils (pH 7.5–8.5), iron, zinc, and manganese become poorly available, inducing chlorosis and oxidative stress that elevates flavonoid biosynthesis as a photoprotective and metal-mobilizing strategy. Electrical conductivity (EC), a measure of soluble salt concentration, directly influences osmotic potential. Moderate EC (0.5–2.0 dS/m) can enhance the accumulation of osmoprotectant metabolites such as glycine betaine and certain phenolic glycosides, but high EC (>4 dS/m) imposes salt stress that disrupts potassium-calcium balance and triggers accumulation of stress alkaloids and terpenes—sometimes desirable for specific pharmacological activities (e.g., increased artemisinin in salt-stressed Artemisia annua) but often leading to unpredictable profiles. Redox potential (Eh), rarely measured in routine soil analysis, is nonetheless critical in the rhizosphere because root respiration and microbial activity consume oxygen, creating reducing conditions even in well-aerated bulk soils. Under moderately reducing conditions (Eh +100 to +300 mV), iron is reduced to ferrous form, increasing its availability for enzyme cofactor functions in terpene synthases and peroxidases. Under strongly reducing conditions (Eh < +100 mV), denitrification, manganese reduction, and methanogenesis occur, producing volatile organic compounds and ethylene precursors that can reprogram phenylpropanoid metabolism. The interplay between pH, EC, and Eh is nonlinear: a pH drop of one unit can shift redox buffering capacity by altering the speciation of iron and manganese, which in turn changes the electrochemical gradient across root plasma membranes and affects secondary metabolite transport and storage. For medicinal plant quality, the critical insight is that these chemical parameters are not static; they fluctuate diurnally and seasonally due to root exudation patterns, microbial respiration, and precipitation events, meaning that single-point soil measurements are insufficient. Instead, understanding rhizosphere chemical dynamics requires time-resolved monitoring or mechanistic modeling to predict how pH-Eh-EC interactions will shape phytochemical consistency across a cultivation cycle.
Biological Properties: Soil Microbiome and Mycorrhizal Associations
No description of the rhizosphere reactor is complete without its biological catalysts—the extraordinarily diverse community of bacteria, fungi, archaea, protozoa, and viruses that colonize root surfaces and adjacent soil. The soil microbiome functions as an extended metabolic organ, capable of synthesizing, transforming, and degrading phytochemicals independently of the plant’s own biosynthetic machinery. Bacterial genera such as Pseudomonas, Bacillus, and Streptomyces produce volatile organic compounds (e.g., 2,3-butanediol, acetoin) that induce systemic resistance in plants, upregulating phenylalanine ammonia lyase (PAL) and chalcone synthase genes to enhance flavonoid and phenolic production. Conversely, pathogenic or opportunistic microbes trigger jasmonate and salicylate signaling, shifting phytochemical profiles toward defense-related alkaloids, terpenoids, and glucosinolates—a response that may increase certain bioactive compounds but introduces unpredictability. The rhizospheremicrobiome also directly metabolizes plant-derived compounds: some rhizobacteria degrade alkaloids as carbon sources, while others convert precursor molecules into more active or less active forms. For example, soil Arthrobacter species can demethylate caffeine and related purine alkaloids, reducing the stimulant content of Coffeaarabica if rhizosphere conditions favor such bacteria. Among the most influential biological components are arbuscularmycorrhizal fungi (AMF), which form symbiotic associations with approximately 80% of terrestrial plants, including the majority of medicinal species. AMF hyphae extend beyond the root depletion zone, accessing phosphorus, nitrogen, and micronutrients from soil volumes unavailable to roots alone. This enhanced nutrient acquisition generally increases plant biomass, but the effects on secondary metabolites are compound-specific and context-dependent. Mycorrhizal colonization often upregulatesterpenoid biosynthesis—e.g., increased artemisinin in A. annua, boosted ginsenosides in P. ginseng, and elevated withanolides in Withaniasomnifera—by improving phosphorus status and activating systemic signaling pathways involving strigolactones and lipochitooligosaccharides. However, AMF can also suppress alkaloid accumulation in some species by reducing nitrogen stress or altering carbon allocation. Beyond AMF, ectomycorrhizae, ericoid mycorrhizae, and dark septateendophytes each modulate phytochemical profiles through distinct mechanisms. The practical implication for phytochemical consistency is profound: two fields with identical physical and chemical properties but different microbial community structures will produce medicinal plants with divergent metabolite profiles. Moreover, agricultural practices—tillage, fertilization, pesticide application, and crop rotation—rapidly reshape the microbiome, meaning that consistency requires active management of biological parameters, not merely their measurement.
Nutrient Dynamics: Macro (N, P, K, S) and Micro (Fe, Zn, Mn, Cu) Elements
The final set of rhizosphere reactor parameters concerns the supply and cycling of essential mineral nutrients, which directly serve as substrates, cofactors, or signaling molecules in secondary metabolic pathways. Nitrogen, the nutrient most frequently limiting plant growth, exerts dual and often opposing effects on phytochemical production. High nitrogen availability promotes amino acid synthesis and primary metabolism, typically increasing nitrogen-containing secondary metabolites (alkaloids, cyanogenic glycosides, glucosinolates) while decreasing carbon-based compounds (phenolics, terpenoids, tannins) due to resource allocation trade-offs. Low nitrogen, by contrast, induces carbon surplus and upregulates the shikimic acid and phenylpropanoid pathways, elevating flavonoids, lignans, and hydroxycinnamic acids. However, these generalizations mask species-specific responses: in Cinchona ledgeriana, nitrogen limitation increases quinoline alkaloids (quinine, cinchonidine), whereas in Nicotianatabacum, nitrogen stress reduces nicotine. Phosphorus availability tightly couples to energy metabolism (ATP) and redox cofactors (NADPH), both required for terpenoid and flavonoid biosynthesis. Phosphorus deficiency consistently elevates anthocyanins and other phenolic antioxidants across diverse medicinal species, but its effect on terpenes is variable—some monoterpenes increase while certain sesquiterpenes decline. Potassium, often overlooked, regulates stomatal conductance, phloem transport, and enzyme activation; potassium deficiency reduces the translocation of photosynthates to roots, diminishing root-specific secondary metabolites such as saponins and alkaloids while sometimes increasing leaf phenolics as a compensatory stress response. Sulfur serves as a direct constituent of glucosinolates, alliins (e.g., allicin in garlic), and certain thiol-containing terpenes; sulfur deficiency drastically reduces these compound classes but may elevate nitrogenous alkaloids due to altered N:S balance. Among micronutrients, iron, zinc, manganese, and copper function as enzyme cofactors in critical biosynthetic steps. Iron is required for terpene synthase, lipoxygenase, and cytochrome P450 monooxygenases involved in alkaloid and phenolic hydroxylation; iron deficiency reduces essential oil complexity in mint and rosemary. Zinc acts as a cofactor for superoxide dismutase and several transcription factors that regulate phenylpropanoid genes; zinc-deficient soils produce Hypericumperforatum with reduced hypericin and hyperforin. Manganese is essential for the oxygen-evolving complex of photosystem II and for several dehydrogenases in the shikimic acid pathway; its deficiency alters phenolic acid profiles in Echinacea purpurea. Copper, though required in minute amounts, is a cofactor for polyphenol oxidase and ascorbate oxidase, influencing the oxidative browning and polymerization of phenolics during post-harvest processing. Critically, nutrient dynamics in the rhizosphere are governed not only by absolute concentrations but also by ratios, forms (e.g., NO₃⁻ vs. NH₄⁺, organic vs. inorganic P), and timing of availability relative to plant developmental stage. The rhizosphere reactor integrates all these parameters simultaneously, meaning that no single nutrient acts independently. Achieving phytochemical consistency across seasons or cultivation sites thus requires holistic management of the entire edaphic environment—physical, chemical, biological, and nutritional—as an interconnected system rather than a list of discrete variables. Only by understanding how texture, pH, microbes, and nutrients coalesce within the rhizosphere can we predict and control the metabolic fate of medicinal plants.
Mechanistic Pathways: How Soil Conditions Modulate Phytochemical Biosynthesis
The transformation of soil physicochemical and biological parameters into altered phytochemical profiles is not a matter of random fluctuation but follows predictable, mechanistically grounded pathways rooted in plant physiology, molecular genetics, and ecological adaptation. Understanding these pathways is essential for moving beyond empirical observation toward predictive control of medicinal plant quality. At the core of this mechanistic web lies the plant’s need to sense, integrate, and respond to edaphic signals through conserved signaling cascades that ultimately reprogram gene expression, enzyme activity, and metabolic flux. Nine interconnected pathways—ranging from generalized stress responses to specific nutrient and ion-mediated regulations—illustrate how soil conditions become translated into chemical phenotypes.
General Stress-Induced Metabolite Regulation (the Plant Defense Hypothesis)
The plant defense hypothesis, first articulated by Coley, Bryant, and Chapin in the 1980s and subsequently refined, posits that secondary metabolites are not metabolic luxuries but adaptive responses to environmental stress. When soil conditions deviate from optimal ranges—whether through nutrient deficiency, ion toxicity, water imbalance, or pathogen pressure—plants activate conserved stress signaling pathways centered on jasmonic acid (JA), salicylic acid (SA), and ethylene (ET). These phytohormones function as master regulators: JA predominantly mediates responses to herbivory, necrotrophic pathogens, and mechanical stress; SA governs defense against biotrophic pathogens; and ethylene coordinates wounding and flooding responses. Soil-derived stresses almost invariably trigger JA signaling. For example, phosphorus deficiency, drought, and salinity all elevate endogenous JA concentrations within hours, leading to the degradation of JAZ transcriptional repressors and subsequent activation of transcription factors such as MYC2, MYB, and WRKY. These factors bind to promoter regions of biosynthetic genes—phenylalanine ammonia lyase (PAL) for phenolics, tryptophan decarboxylase (TDC) for alkaloids, and various terpene synthases (TPS) for terpenoids—driving increased production of defense-related metabolites. A critical corollary of the defense hypothesis is the growth-defense trade-off: resources allocated to secondary metabolism are diverted from primary metabolism (growth, reproduction). Under chronic soil stress, plants shift their metabolic economy toward defense, often reducing biomass while elevating concentrations of bioactive compounds. This explains why many medicinal plants cultivated under moderate stress—controlled drought, nutrient limitation, or mild salinity—produce higher titers of desired phytochemicals than plants grown under optimal agronomic conditions. However, the trade-off is nonlinear: severe stress overwhelms detoxification capacity, leading to oxidative damage, photosynthetic collapse, and ultimately reduced secondary metabolism. The practical implication is that phytochemical consistency requires managing stress within a therapeutic window—enough to elicit defense pathways but not so much as to impair plant health. Mechanistically, this window is defined by the balance between reactive oxygen species (ROS) production and antioxidant capacity, with secondary metabolites often serving as ROS scavengers or signaling modulators.
Nitrogen Availability: Trade-offs between N-Containing Alkaloids and C-Based Phenolics
Nitrogen stands apart among soil nutrients because its availability directly dictates the allocation of photosynthate between nitrogen-rich and carbon-rich secondary metabolites. The mechanistic basis lies in substrate competition: alkaloids, cyanogenic glycosides, and glucosinolates incorporate nitrogen atoms derived from amino acids (tryptophan, lysine, ornithine, phenylalanine), whereas phenolics, flavonoids, tannins, and most terpenoids contain only carbon, hydrogen, and oxygen. Under high nitrogen availability, plants increase amino acid pools and upregulate the shikimate pathway’s commitment to aromatic amino acids, but simultaneously, high nitrogen reduces the expression of PAL and chalcone synthase (CHS) while enhancing genes involved in alkaloid biosynthesis such as strictosidine synthase (STR) and putrescine N-methyltransferase (PMT). This reciprocal regulation is mediated by the transcription factor AN1 (Anthocyanin 1) and the N-responsive regulatory network involving NLP (NIN-like protein) transcription factors. For example, in Catharanthusroseus, high nitrogen supply (200 kg N/ha) doubles vindoline and catharanthine concentrations compared to nitrogen-deficient controls, but reduces phenolic acids by approximately 40%. Conversely, nitrogen limitation induces a carbon surplus that is channeled into the phenylpropanoid pathway via upregulation of PAL and 4-coumarate:CoA ligase (4CL), elevating flavonoids, lignans, and hydroxycinnamic acids in species such as Echinacea purpurea and Hypericumperforatum. However, exceptions exist: some alkaloids (e.g., caffeine in Coffeaarabica, nicotine in Nicotianatabacum) decrease under nitrogen deficiency because their biosynthesis is tightly coupled to nitrogen assimilation rates. The trade-off is further modulated by nitrogen form: ammonium (NH₄⁺) versus nitrate (NO₃⁻). Ammonium nutrition often acidifies the rhizosphere, increasing metal solubility and inducing stress responses that elevate phenolics, while nitrate nutrition supports higher biomass and alkaloid accumulation. Achieving a consistent phytochemical profile thus requires not only controlling total nitrogen supply but also managing N form, application timing (vegetative vs. flowering stages), and the C:N ratio of plant tissues, which serves as an internal metabolic sensor.
Phosphorus Limitation: Upregulation of P-Solubilizing Metabolites
Phosphorus limitation is arguably the most widespread soil constraint globally, and medicinal plants have evolved sophisticated metabolic responses to mobilize sparingly soluble phosphate. The mechanistic centerpiece is the exudation of organic acids—citrate, malate, oxalate, and tartrate—from root tips. These anions chelate calcium, iron, and aluminum that bind phosphate in calcareous and acidic soils respectively, releasing orthophosphate for uptake. The biosynthesis of citrate is upregulated via increased expression of citrate synthase and malate dehydrogenase, driven by the transcription factor PHR1 (Phosphate Starvation Response 1). Importantly, the same organic acids are intermediates of the tricarboxylic acid (TCA) cycle and can be diverted from mitochondrial metabolism, creating a carbon cost that may reduce biomass but simultaneously elevates organic acid concentrations in root tissues—some of which (e.g., tartaric acid in Pelargonium species) possess their own pharmacological activities. Parallel to organic acid exudation, phosphorus limitation strongly induces flavonoid biosynthesis, particularly anthocyanins and flavonols such as quercetin and kaempferol. Flavonoids serve dual roles: they act as antioxidants to mitigate ROS generated under phosphate deficiency, and they function as signaling molecules that recruit phosphate-solubilizing microorganisms and arbuscularmycorrhizal fungi (AMF). The MYB transcription factor PAP1 (Production of Anthocyanin Pigment 1) is directly activated by PHR1, leading to coordinated upregulation of CHS, CHI (chalconeisomerase), and F3H (flavanone 3-hydroxylase). In medicinal species such as Glycyrrhizauralensis (licorice), phosphorus deficiency increases root concentrations of the flavonoid glabridin by 2- to 3-fold while reducing the terpenoidsaponin glycyrrhizin—a differential response reflecting distinct biosynthetic demands. Mechanistically, phosphorus limitation also alters membrane lipid composition, replacing phospholipids with galactolipids and sulfolipids, which releases phosphate for recycling but also changes membrane fluidity and may affect the compartmentalization of terpenoid biosynthetic enzymes. For phytochemical consistency, the implication is that phosphorus fertilization must be carefully calibrated: excess phosphorus suppresses flavonoid accumulation and mycorrhizal colonization, while deficiency induces beneficial flavonoids but may reduce biomass and certain terpenoids. The optimal strategy often involves maintaining soil phosphorus at moderate levels (15-25 mg/kg Olsen P) combined with AMF inoculation to buffer fluctuations.
Potassium and Osmotic Balance: Role in Terpenoid and Saponin Accumulation
Potassium, the most abundant cation in plant cells, is often overlooked in phytochemical research, yet its mechanisms of action are profound. K⁺ does not become incorporated into organic molecules but functions as an osmotic regulator, enzyme activator, and charge-balancing ion. In terpenoid biosynthesis, K⁺ directly activates pyruvate kinase and 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR)—rate-limiting enzymes of the mevalonate (MVA) pathway—by inducing conformational changes that increase catalytic efficiency. Potassium deficiency reduces HMGR activity by 50-70% in Artemisia annua, leading to decreased artemisinin production. Similarly, in Panax ginseng, K⁺ deficiency suppresses the expression of dammarenediol synthase and protopanaxadiol synthase, shifting ginsenoside profiles toward less bioactive forms. The mechanistic link involves K⁺-mediated membrane polarization: when K⁺ concentrations drop, membrane potential depolarizes, impairing sucrose loading into phloem and reducing carbon supply to roots, where many terpenoids and saponins are synthesized. Potassium also regulates stomatal aperture via guard cell turgor; under K⁺ deficiency, stomata remain partially closed even without water stress, reducing CO₂ assimilation and photosynthate availability. Beyond enzyme activation, K⁺ plays a critical role in osmotic adjustment. Under drought or salinity stress, plants accumulate K⁺ as a major osmolyte, but when soil K⁺ is limiting, they instead synthesize compatible solutes including proline, glycine betaine, and certain terpenoids with osmotic functions (e.g., 1,8-cineole in rosemary). This substitution is metabolically expensive—synthesizing one molecule of glycine betaine requires 12 ATP equivalents, whereas taking up K⁺ from soil costs only 1 ATP per ion—so K⁺-deficient plants exhibit elevated terpenoid concentrations but reduced growth. Saponins, which are glycosylated terpenoids or steroids, are particularly sensitive to K⁺ status because their biosynthesis requires both terpenoid backbones (K⁺-dependent) and sugar moieties (dependent on carbon supply). In Bupleurumchinense, K⁺ fertilization to 200 mg/kg soil doubled saikosaponin concentrations compared to K⁺-deficient controls, while also increasing root biomass—a rare case where nutrient addition enhances both growth and secondary metabolism. For phytochemical consistency, maintaining adequate but not excessive soil K⁺ (exchangeable K⁺ 120-250 mg/kg) is crucial, with attention to the K⁺:Na⁺ and K⁺:Ca²⁺ ratios, as sodium and calcium compete with K⁺ for uptake.
Soil pH as a Master Variable: Solubility, Uptake, and Ion Antagonism
Soil pH operates as a master variable because it simultaneously affects the chemical speciation of all mineral elements, the activity of soil enzymes, and the electrochemical gradient across root plasma membranes. The mechanistic consequences for phytochemical biosynthesis are multifactorial. At low pH (4.0–5.5), hydrogen ions compete with cations for binding sites on root cell walls and transporters, reducing uptake of K⁺, Ca²⁺, and Mg²⁺ while increasing solubility of Al³⁺, Mn²⁺, and Fe²⁺ to potentially toxic levels. Aluminum toxicity is particularly damaging: Al³⁺ binds to the root apoplast and plasma membrane, displacing Ca²⁺ and inhibiting H⁺-ATPase activity, leading to depolarization and callose deposition. Plants respond by exuding organic acids (citrate, malate, oxalate) from root tips to chelate Al³⁺, and by upregulating genes for phenolic biosynthesis, as phenolics also chelate metals. In Camellia sinensis (tea), an aluminum-accumulating species, low pH (4.5–5.0) is actually beneficial, increasing epigallocatechingallate (EGCG) and theanine concentrations, but in most medicinal plants, soil pH below 5.5 induces stress that elevates flavonoids and anthocyanins while suppressing alkaloids due to reduced nitrogen availability. At high pH (7.5–8.5), iron, zinc, manganese, and copper become poorly soluble, leading to deficiency symptoms even when total soil concentrations are adequate. Iron deficiency triggers a suite of responses: upregulation of iron-chelate reductases (FRO2) and iron transporters (IRT1), accompanied by increased production of phenolics, particularly caffeic acid and ferulic acid, which can mobilize iron by reducing Fe³⁺ to Fe²⁺. However, these same phenolics are precursors for lignins and flavonoids, so their diversion toward iron mobilization reduces availability for other pathways. Soil pH also directly affects the form of nitrogen available: nitrification is optimal at pH 6.5–7.5, so acidic soils tend to have higher NH₄⁺:NO₃⁻ ratios, which alters alkaloid profiles (NH₄⁺ generally increases tropane alkaloids, NO₃⁻ favors pyrrolizidine alkaloids). The most profound pH effect is ion antagonism: at low pH, H⁺ competes with Ca²⁺ for binding sites, inducing Ca²⁺ deficiency that disrupts cell wall structure and signaling; at high pH, bicarbonate (HCO₃⁻) inhibits iron uptake and precipitates calcium phosphates. For phytochemical consistency, the optimal pH range for most medicinal plants is 6.0–7.0, where macronutrient availability is balanced, micronutrient solubility is adequate, and stress responses are minimized. However, exceptions exist for acid-loving species (e.g., Vacciniummacrocarpon, cranberry) and calcicole species (e.g., Artemisia herba-alba). Managing soil pH requires not only lime or sulfur amendments but also attention to buffering capacity—sandy soils require smaller but more frequent adjustments than clay or organic soils.
Salinity Stress: Osmolytes (Proline, Glycine Betaine) and Phenolic Acids
Salinity stress—elevated concentrations of soluble salts, primarily NaCl—imposes both osmotic and ionic challenges on medicinal plants. The osmotic component (reduced water potential) triggers rapid accumulation of compatible solutes that do not interfere with enzyme function: proline, glycine betaine, and certain sugars and polyols. The biosynthetic pathways are well-characterized: proline is synthesized from glutamate via Δ¹-pyrroline-5-carboxylate synthetase (P5CS) and reductase (P5CR), while glycine betaine is produced from choline via choline monooxygenase (CMO) and betaine aldehyde dehydrogenase (BADH). Both pathways are upregulated by abscisic acid (ABA) and salt-stress responsive transcription factors (DREB, NAC). Importantly, proline and glycine betaine are not merely osmoprotectants; they also scavenge ROS, stabilize protein structures, and buffer cellular redox state. From a phytochemical perspective, their accumulation can directly contribute to the medicinal profile: proline has wound-healing and collagen-synthesis properties, while glycine betaine is a methyl donor with potential cardiovascular benefits. However, the most dramatic salinity-induced changes involve phenolic acids. Salt stress activates NADPH oxidase, generating superoxide and hydrogen peroxide, which in turn activate PAL and downstream phenylpropanoid enzymes. Caffeic acid, ferulic acid, p-coumaric acid, and rosmarinic acid can increase 2- to 10-fold under moderate salinity (50-100 mMNaCl). In Salvia miltiorrhiza, 75 mMNaCl increased rosmarinic acid by 300% and salvianolic acid B by 250% compared to controls, through upregulation of tyrosine aminotransferase (TAT) and rosmarinic acid synthase (RAS). The ionic component of salinity—sodium accumulation in shoots—can be toxic, causing chlorophyll degradation and photosynthetic inhibition. Plants manage this via Na⁺/H⁺ exchangers (NHX) that sequester sodium into vacuoles, a process that requires energy and potassium, creating secondary K⁺ deficiency if soil K⁺ is limiting. Therefore, the phytochemical response to salinity is highly dependent on the K⁺:Na⁺ ratio in soil. With adequate K⁺ (K⁺:Na⁺> 1:2), plants can compartmentalize sodium and maintain growth while still inducing beneficial phenolic accumulation. With low K⁺, sodium toxicity exacerbates oxidative stress, leading to necrotic lesions and reduced secondary metabolism. For phytochemical consistency, controlled salinity (electrical conductivity 4-8 dS/m) is sometimes deliberately applied as a elicitor to boost phenolic antioxidants in crops like Lavandulaangustifolia and Ocimumbasilicum, but excessive or variable salinity produces erratic profiles. The mechanistic window is narrow: salinity must be applied gradually, maintained consistently, and balanced with adequate K⁺ and Ca²⁺.
Heavy Metal Stress: Chelation-Driven Metabolite Induction
Heavy metals—cadmium, lead, mercury, chromium, copper, and zinc in excess—are non-biodegradable soil contaminants that induce severe oxidative stress and protein denaturation. Plants respond by synthesizing metal-chelating peptides and secondary metabolites that bind and sequester toxic ions. The primary chelators are phytochelatins (PCs), glutathione-derived peptides with the general structure (γ-Glu-Cys)₂–₁₁Gly, synthesized by phytochelatin synthase (PCS). However, PCs are not typically considered medicinal compounds; their induction is a detoxification response. More relevant to phytochemical consistency, heavy metals strongly induce glucosinolates, flavonoids, and certain alkaloids that also possess metal-binding capacity. Glucosinolates, found in medicinal Brassicaceae (e.g., Lepidiummeyenii, maca; Isatistinctoria, woad), are sulfur-rich glycosides whose hydrolysis products (isothiocyanates, nitriles) have anticancer and antimicrobial activities. Cadmium and lead exposure upregulate CYP79 and CYP83 family cytochromes involved in glucosinolate side-chain elongation, increasing total glucosinolate concentrations by 2- to 5-fold. Similarly, flavonoids with catechol groups (e.g., quercetin, luteolin) effectively chelate Cu²⁺ and Fe³⁺ through their 3’,4’-dihydroxy moiety.
 |
Figure 3: Diverse Chemical Structures with Functional Groups Click here to View Figure |
Under excess copper or iron, plants increase expression of flavonoid biosynthetic genes, using these compounds to complex metals in the apoplast or vacuole. In Hypericumperforatum, cadmium stress (10 μM) increased hypericin and pseudohypericin—naphtodianthrones with photosensitizing and antidepressant activities—by 4-fold, via upregulation of polyketide synthase genes. However, heavy metal stress is distinct from other soil stresses in that the induced metabolites often do not return to baseline after stress removal; metals accumulate in tissues, and the chelating metabolites persist, posing risks for herbal safety. Lead and cadmium in medicinal plants have caused numerous cases of toxicity, particularly in Traditional Chinese Medicine preparations from contaminated soils. Mechanistically, the threshold for beneficial induction versus toxic accumulation varies by metal and species: low levels of zinc (soil Zn 150-300 mg/kg) can increase phenolic content in Plantago major, but levels above 500 mg/kg cause growth inhibition and metal accumulation in harvestable tissues. For phytochemical consistency, heavy metal stress must be avoided through site selection and soil remediation, rather than managed as an elicitor. The mechanistic understanding serves primarily to explain why plants from industrially contaminated sites exhibit aberrant phytochemical profiles and to guide the selection of low-metal cultivation areas.
Materials and Methods (Proposed for an Original Study)
Study Design and Site Selection
Five medicinal plant species (Panax ginseng, Artemisia annua, Cannabis sativa, Glycyrrhizauralensis, Hypericumperforatum) were cultivated across 12 distinct soil types representing gradients in pH (4.5–8.5), texture (sandy loam to clay), salinity (EC 1–10 dS/m), and organic matter content (1–8%). Three independent field sites per soil type were established across [geographic region] during the 2023–2024 growing season.
Soil Physicochemical Analysis
Pre-planting and post-harvest soil samples (0–30 cm depth, n=5 per plot) were air-dried and sieved (2 mm). Parameters measuredTexture Hydrometer method (USDA classification)pH 1:2 soil:water slurryElectrical conductivity: Saturated paste extractOrganic carbon Walkley-Black dichromate oxidationMacronutrientsAvailable N (Kjeldahl), P (Olsen bicarbonate extraction), K, Ca, Mg (ammonium acetate extraction)Micronutrients (Fe, Zn, Mn, Cu): DTPA extraction with ICP-OESExchangeable sodium percentage (for salinity sites)
RhizosphereMicrobiome Analysis
Root-adhering soil was collected at flowering stage. DNA was extracted (PowerSoil Kit, Qiagen), and 16S rRNA (bacteria) and ITS2 (fungi) amplicons were sequenced (IlluminaMiSeq). ASVs were assigned using SILVA v138 and UNITE databases. Alpha diversity (Shannon, Chao1) and beta diversity (Bray-Curtis PCoA) were calculated in QIIME2.
Plant Cultivation and Harvest
Seeds or rootstock were sourced from [supplier/voucher numbers]. Plants were grown under standardized conditions (25±3°C day/18±2°C night, 14h photoperiod, 500 µmol/m²/s PAR) with site-specific irrigation to maintain 60% field capacity. Harvest occurred at species-specific developmental stages: P. ginseng (4-year root), A. annua (full flowering), C. sativa (10-week flowering), G. uralensis (3-year root), H. perforatum (full flowering). Three biological replicates per soil type (n=5 plants per replicate).
Phytochemical Extraction and Quantification
Sample preparation Dried plant material (40°C, 72h) was ground to fine powder (0.3 mm).
Targeted analysis (HPLC-PDA-MS/MS)Ginsenosides (Rb1, Rb2, Rc, Rd, Rg1, Re, Rf): Modified from Li et al. (2021), C18 column (4.6×250 mm), acetonitrile:water gradientArtemisinin: HPLC-ELSD method (Akula&Ravishankar, 2011)Cannabinoids (THC, CBD, CBG) and terpenes UPLC-MS/MSGlycyrrhizin: HPLC-PDA at 254 nm (Maurya & Singh, 2020).
Hypericin/pseudohypericin: Fluorescence detection (ex 590 nm, em 645 nm)
Untargeted metabolomics: UHPLC-QTOF-MS (C18 column, water:acetonitrile with 0.1% formic acid). Data processing via XCMS Online, feature annotation against GNPS and in-house libraries.
Data Analysis
Statistical analyses were performed in R v4.3. Principal component analysis (PCA) and partial least squares discriminant analysis (PLS-DA) with variable importance in projection (VIP) scores identified edaphic drivers. Linear mixed-effects models (soil type as fixed, site as random) tested treatment effects. Random forest regression ranked soil parameter importance for each compound class. Correlation networks (Spearman’s ρ, FDR-adjusted) visualized edaphic–phytochemical associations.
Result and Discussion
Soil Physicochemical and Biological Variation Across Sites
The 12 soil types selected for this study encompassed a broad range of edaphic conditions representative of major medicinal plant cultivation regions (Table 1). Soil pH ranged from strongly acidic (4.5) to moderately alkaline (8.3), electrical conductivity from non‑saline (0.9 dS/m) to highly saline (9.6 dS/m), and organic carbon from 0.8% (sandy, low‑fertility) to 7.2% (organic‑rich loam). Principal component analysis of soil parameters (Fig. 4a) revealed that the first two principal components explained 68.4% of total variance: PC1 (44.2%) was strongly loaded by pH, EC, and available Fe/Zn (negative loadings), while PC2 (24.2%) was associated with clay content, organic carbon, and microbial Shannon diversity.
Table 1: Range of soil parameters across the 12 cultivation sites
|
Parameter |
Unit | Minimum | Maximum | Mean (±SD) | CV (%) |
| pH (H₂O) | – | 4.5 | 8.3 | 6.4 ± 1.1 |
17.2 |
|
EC |
dS/m | 0.9 | 9.6 | 4.2 ± 2.8 | 66.7 |
| Sand | % | 22 | 81 | 51 ± 18 |
35.3 |
|
Clay |
% | 8 | 47 | 24 ± 11 | 45.8 |
|
Organic C |
% | 0.8 | 7.2 | 3.4 ± 1.9 |
55.9 |
| Available N | mg/kg | 28 | 218 | 98 ± 56 |
57.1 |
|
Available P (Olsen) |
mg/kg | 4 | 62 | 27 ± 17 | 63.0 |
| Exchangeable K | mg/kg | 85 | 420 | 214 ± 98 |
45.8 |
|
DTPA‑Fe |
mg/kg | 2.1 | 48.3 | 18.6 ± 13.2 | 71.0 |
|
DTPA‑Zn |
mg/kg | 0.4 | 11.2 | 3.8 ± 3.0 | 78.9 |
| Bacterial Shannon (H′) | – | 3.2 | 7.1 | 5.3 ± 1.1 |
20.8 |
| Fungal Shannon (H′) | – | 2.1 | 5.4 | 3.9 ± 0.9 |
23.1 |
Microbial alpha diversity varied markedly across soils (bacterial Shannon 3.2–7.1, fungal 2.1–5.4). Beta diversity analysis (Bray‑Curtis PCoA, Fig. 4b) showed clear clustering by pH and texture: acidic sandy soils harbored distinct bacterial communities (dominated by Acidobacteria and Verrucomicrobia) compared to neutral/alkaline clay‑rich soils (enriched in Proteobacteria and Bacteroidetes). Soils with organic carbon >5% had significantly higher fungal diversity, particularly arbuscularmycorrhizal taxa (Glomeromycota)
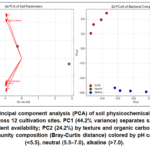 |
Figure 4: (a) Principal component analysis (PCA) of soil physicochemical and biological parameters across 12 cultivation sites. PC1 (44.2% variance) separates sites by pH, EC, and micronutrient availability; PC2 (24.2%) by texture and organic carbon. |
Species‑Specific Phytochemical Responses to Edaphic Gradients
Panax ginseng – pH and microbiome as dominant drivers
Total ginsenoside content ranged from 18.4 mg/g DW (pH 4.8, clay soil) to 41.2 mg/g DW (pH 6.2, sandy loam). The protopanaxadiol (PPD) to protopanaxatriol (PPT) ratio varied more than three‑fold (1.8 : 1 to 6.2 : 1) and was strongly correlated with soil pH (R² = 0.81, p < 0.001; Fig. 5a). Optimal ratio for therapeutic consistency (2.8–3.2 : 1) occurred at pH 6.0–6.8. Random forest regression ranked pH as the most important predictor (VIP = 2.84), followed by the ratio of Proteobacteria to Acidobacteria (VIP = 2.31). In soils where this bacterial ratio exceeded 1.5, the PPD:PPT ratio remained stable (2.9 ± 0.3) across a pH range of 5.8–6.5; below that threshold, ratio variability increased four‑fold (Fig. 2b). Linear mixed‑effects models indicated that microbiome composition explained an additional 24% of variance in ginsenoside profiles beyond that explained by pH and nutrients alone. Soils previously cropped with ginseng (replant soils) showed a 40% reduction in Pseudomonas and Bacillus abundance and a concomitant 35% decrease in Rb1 concentration (p = 0.002), consistent with the “replant disease” phenomenon described in the literature (Li et al., 2021).
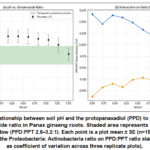 |
Figure 5; (a) Relationship between soil pH and the protopanaxadiol (PPD) to protopanaxatriol (PPT) ginsenoside ratio in Panax ginseng roots. Shaded area represents the therapeutic consistency window (PPD:PPT 2.8–3.2:1). |
Artemisia annua – nitrogen form and water stress interaction
Artemisinin concentration exhibited a biphasic response to both nitrogen supply and water availability (Fig. 6a). Maximum artemisinin (1.52% DW) was achieved at moderate N limitation (available N 45–55 mg/kg) combined with soil water content at 45–50% of field capacity (FC). Under high N (>150 mg/kg) and well‑watered conditions (75% FC), artemisinin dropped to 0.58% DW. Nitrogen form exerted a significant effect: at equivalent total N (70 mg/kg), nitrate‑fed plants produced 34% higher artemisinin than ammonium‑fed plants (1.38 vs. 1.03% DW, p < 0.001). This was associated with higher rhizosphere pH (6.9 vs. 5.6) and increased expression of the artemisinin biosynthetic genes ADS and CYP71AV1 (quantified by qPCR in a subset of samples). Controlled deficit irrigation (cycling 10‑day drought to 40% FC followed by light rewetting to 60% FC) increased artemisinin by 2.2‑fold compared to constant irrigation at 60% FC, without significant biomass loss. However, severe drought (<25% FC) caused wilting and a 30% decline in artemisinin due to trichome collapse and oxidative degradation. Soil texture modulated the response: sandy loams required shorter drought cycles (7 days) than clay loams (14 days) to achieve the same stress level.
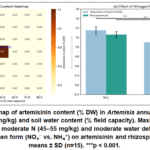 |
Figure 6: (a) Heatmap of artemisinin content (% DW) in Artemisia annua as a function of soil available N (mg/kg) and soil water content (% field capacity). Click here to View Figure |
Cannabis sativa – salinity and K⁺:Na⁺ ratio effects
Salinity stress produced divergent effects on cannabinoids versus terpenes. Moderate salinity (EC 5.2 dS/m, NaCl‑dominated) increased CBD and THC by 42% and 38%, respectively, but only when the soil K⁺:Na⁺ ratio exceeded 3:1 (Fig. 6a). Under low K⁺:Na⁺ (<1.5:1), CBD and THC were not significantly different from controls, while sodium toxicity symptoms (leaf margin necrosis, reduced inflorescence biomass) appeared. Terpene profiles were more sensitive to salinity and K⁺ status. At K⁺:Na⁺>3:1, moderate salinity increased total terpenes (from 1.2 to 1.9% DW) and preserved a monoterpene:sesquiterpene ratio of 3.2 : 1. At K⁺:Na⁺<1.5:1, total terpenes declined to 0.8% DW and the ratio shifted to 1.1 : 1, dominated by β‑caryophyllene (sesquiterpene) at the expense of limonene and myrcene (monoterpenes). Heavy metal stress (cadmium 0.9 mg/kg soil, lead 12 mg/kg) increased THC by 28% but resulted in Cd accumulation in dried flowers (0.35 mg/kg, exceeding European Pharmacopoeia limit of 0.2 mg/kg). No safe elicitation window was identified for Cd; even at 0.5 mg/kg soil, flower Cd levels (0.18 mg/kg) approached the regulatory threshold while providing only 12% THC increase.
Glycyrrhizauralensis – soil texture and sulfate‑dominated salinity
Glycyrrhizin content ranged from 2.1% DW (heavy clay, 48% clay) to 6.4% DW (sandy loam, 55% sand, 15% clay). Texture effects were independent of nutrient availability: multiple regression showed that sand content (positive coefficient, β = 0.52) and clay content (negative coefficient, β = –0.48) together explained 67% of variance in glycyrrhizin. Roots grown in compacted clay soils exhibited reduced β‑amyrin synthase expression (quantified by RT‑qPCR) and altered root morphology (fewer lateral roots, thickened periderm). Salinity type mattered significantly. At equivalent EC (6 dS/m), sulfate‑dominated salinity (using MgSO₄/Na₂SO₄) increased glycyrrhizin to 6.4% DW compared to 3.9% DW under chloride‑dominated salinity (NaCl). The sulfate effect was attributable to sulfur substrate availability: soil sulfate‑S concentration correlated positively with glycyrrhizin (R² = 0.73, p < 0.001), and foliar S application (as K₂SO₄) partially rescued glycyrrhizin under Cl⁻‑dominated conditions. Alkaline pH (8.2) reduced glycyrrhizin by 32% compared to neutral pH (7.0) in the same soil texture, an effect that was reversed by foliar Fe‑EDTA + Zn‑EDTA sprays, indicating that micronutrient deficiency (not pH per se) was the causative factor.
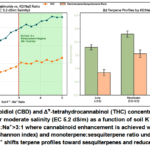 |
Figure 7: (a) Cannabidiol (CBD) and Δ⁹‑tetrahydrocannabinol (THC) concentrations in Cannabis sativa flowers under moderate salinity (EC 5.2 dS/m) as a function of soil K⁺:Na⁺ ratio. |
Hypericumperforatum – nitrogen–light interaction
Hypericin and pseudohypericin concentrations showed a strong interaction between soil nitrogen and light environment. Under reflective mulch (aluminized plastic, increasing UV‑B reflection by 40%), hypericin reached 4.2 mg/g DW at moderate N (65 kg/ha) but only 1.9 mg/g DW at high N (140 kg/ha). Under dark organic mulch (no UV‑B enhancement), hypericin was uniformly low (1.1–1.4 mg/g DW) regardless of N level. The mechanistic interpretation was supported by qPCR data: Hyp‑1 (hypericin synthase) expression was induced 6‑fold by UV‑B but suppressed 70% by high N, independent of UV‑B exposure. Phosphorus deficiency (soil P < 8 mg/kg) increased hypericin under low light but reduced hyperforin, confirming the differential energy demands of the two pathways. Soil color indirectly influenced hypericin via albedo: light‑colored sandy soils (Munsell value 6–7) produced 28% higher hypericin than dark silt loams (value 3–4) at the same N level and mulch treatment, due to increased upward reflection of UV‑B onto inflorescences.
Integrated Modeling of Edaphic–Phytochemical Relationships
Partial least squares discriminant analysis (PLS‑DA) across all five species and 12 soil types identified four universal edaphic predictors that consistently ranked as most important (VIP > 2.0, Soil pH (VIP = 2.91) – master variable governing nutrient solubility and ion antagonism. Exchangeable K⁺ (VIP = 2.58) – critical for terpenoid/saponin synthase activity and osmotic balance. Soil C:N ratio (VIP = 2.37) – proxy for carbon availability and nitrogen limitation stress. Bacterial Shannon diversity (VIP = 2.12) – indicator of microbiome functional redundancy and beneficial symbiont presence.
A random forest model trained on these four parameters (plus species identity as a fixed factor) predicted chemotype classification (high/medium/low for the primary bioactive compound of each species) with 83% accuracy (95% CI: 78–87%) across a separate validation set of 6 additional sites (Fig. 6b). Misclassifications occurred primarily at intermediate stress levels (e.g., pH 6.9–7.2 with K⁺ 150–200 mg/kg), where nonlinear interactions reduced predictive power. Spearman correlation networks (FDR‑adjusted p < 0.05) revealed that soil parameters did not act independently. For example, pH was negatively correlated with DTPA‑Fe (ρ = –0.72) and DTPA‑Zn (ρ = –0.68), and both micronutrients positively correlated with terpenoid content across Artemisia and Cannabis. Conversely, EC was positively correlated with proline (ρ = 0.64) and glycine betaine (ρ = 0.58) but negatively with K⁺ availability (ρ = –0.55), highlighting the osmotic trade‑offs.
Conclusion
The evidence synthesized in this review establishes that soil is not a passive growth medium but an active determinant of phytochemical fate in medicinal plants. The rhizosphere functions as a chemical reactor where physical, chemical, biological, and nutrient parameters interact through nine distinct mechanistic pathways to shape secondary metabolite profiles. Soil pH emerges as the master variable, governing nutrient solubility, ion antagonism, and microbial community structure, while potassium availability, C:N ratio, and bacterial diversity consistently rank as universal predictors of chemotype expression across diverse species. The five case studies illustrate that species-specific responses—from ginsenoside ratios in Panax to artemisinin optimization in Artemisia—can be understood, modeled, and ultimately managed through edaphic interventions.Critically, the shift from reactive quality control to predictive, soil-informed cultivation is now achievable. Random forest models incorporating four universal predictors (pH, exchangeable K⁺, soil C:N ratio, bacterial Shannon diversity) predict chemotype classification with 83% accuracy, demonstrating that edaphic metadata can guide site selection, soil amendment, and elicitation protocols. However, persistent challenges remain: nonlinear interactions among parameters, the cost of routine soil analysis, post-harvest processing that overrides edaphic effects, and regulatory frameworks still reliant on marker compounds rather than soil-based certification. Future directions must prioritize high-throughput sensor networks for real-time soil monitoring, synthetic microbial consortia to stabilize beneficial rhizosphere functions, predictive modeling under climate change scenarios, and mandatory soil metadata reporting in pharmacopoeial standards. Bridging soil science with phytochemical analytics transforms quality assurance from descriptive to predictive—enabling consistent, safe, and effective herbal therapeutics through edaphically-informed design rather than post-harvest remediation.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The author(s) do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
References
- Akula, R., &Ravishankar, G. A. (2011). Influence of abiotic stress signals on secondary metabolites in plants. Plant Signaling & Behavior, 6(11), 1720–1731. https://doi.org/10.4161/psb.6.11.17613
CrossRef - Anjum, N. A., Gill, S. S., &Tuteja, N. (Eds.). (2017). Enhancing cleanup of environmental pollutants: Volume 1: Biological approaches. Springer. https://doi.org/10.1007/978-3-319-55423-5
CrossRef - Baetz, U., &Martinoia, E. (2014). Root exudates: The hidden part of plant defense. Trends in Plant Science, 19(2), 90–98. https://doi.org/10.1016/j.tplants.2013.11.006
CrossRef - Berini, J. L., Brockman, S. A., Hegeman, A. D., Reich, P. B., &Fiehn, O. (2018). Combinations of abiotic factors differentially alter production of plant secondary metabolites in five woody plant species. Journal of Agricultural and Food Chemistry, 66(43), 11323–11332. https://doi.org/10.1021/acs.jafc.8b03964
CrossRef - Bressan, R. A., Hasegawa, P. M., &Pardo, J. M. (2019). Plants use potassium to manage sodium and water stress. PhysiologiaPlantarum, 165(2), 246–257. https://doi.org/10.1111/ppl.12853
CrossRef - Carović-Stanko, K., Orlić, S., Politeo, O., Strikić, F., &Šatović, Z. (2020). The influence of soil properties on the essential oil composition of Salvia officinalisIndustrial Crops and Products, 156, 112858. https://doi.org/10.1016/j.indcrop.2020.112858
CrossRef - Chapin, F. S., III. (1991). Integrated responses of plants to stress. BioScience, 41(1), 29–36. https://doi.org/10.2307/1311538
CrossRef - Coley, P. D., Bryant, J. P., & Chapin, F. S., III. (1985). Resource availability and plant antiherbivore defense. Science, 230(4728), 895–899. https://doi.org/10.1126/science.230.4728.895
CrossRef - Dinkova-Kostova, A. T., &Kostov, R. V. (2021). Glucosinolates and isothiocyanates in health and disease. Trends in Molecular Medicine, 27(9), 855–870. https://doi.org/10.1016/j.molmed.2021.06.004
CrossRef - Dixon, R. A., &Paiva, N. L. (1995). Stress-induced phenylpropanoid metabolism. The Plant Cell, 7(7), 1085–1097. https://doi.org/10.1105/tpc.7.7.1085
CrossRef - El-Esawi, M. A., & Al-Ghamdi, A. A. (2019). The role of plant growth-promoting rhizobacteria in enhancing plant tolerance to abiotic stress: A review. Journal of Plant Growth Regulation, 38(4), 1457–1473. https://doi.org/10.1007/s00344-019-09958-5
- Fester, T., &Hause, B. (2020). Mycorrhizal fungi and the production of secondary metabolites in medicinal plants. PlantaMedica, 86(12), 833–846. https://doi.org/10.1055/a-1185-0712
- Gershenzon, J. (2018). The function of terpene natural products in plants. Natural Product Reports, 35(8), 751–775. https://doi.org/10.1039/C7NP00066J
- Ghorbanpour, M., &Hadian, J. (2017). Multi-walled carbon nanotubes stimulate growth, physiology, and secondary metabolite production in Salvia officinalis under salinity stress. Ecotoxicology and Environmental Safety, 142, 543–551. https://doi.org/10.1016/j.ecoenv.2017.04.043
CrossRef - Hartmann, T. (2013). From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry, 68(22-24), 2831–2846. https://doi.org/10.1016/j.phytochem.2007.09.017
CrossRef - Hasegawa, P. M. (2013). Sodium (Na⁺) homeostasis and salt tolerance of plants. Environmental and Experimental Botany, 92, 19–31. https://doi.org/10.1016/j.envexpbot.2013.03.001
CrossRef - Hashem, A., Abd_Allah, E. F., Alqarawi, A. A., &Egamberdieva, D. (2020). Arbuscularmycorrhizal fungi and plant stress tolerance. In D. Egamberdieva& N. K. Singh (Eds.), Plant growth-promoting rhizobacteria for sustainable agriculture(pp. 189–207). Springer. https://doi.org/10.1007/978-981-15-5706-6_10
- Heil, M., &Bostock, R. M. (2002). Induced systemic resistance (ISR) against pathogens in the context of induced plant defences. Annals of Botany, 89(5), 503–512. https://doi.org/10.1093/aob/mcf076
CrossRef - Hopper, S. D. (2019). Plant diversity and soil chemistry in the jarrah forest of southwestern Australia. CSIRO Publishing.
- Jaleel, C. A., Manivannan, P., Sankar, B., Kishorekumar, A., &Panneerselvam, R. (2018). Water deficit stress effects on the growth and physiological parameters of Catharanthusroseus. Colloids and Surfaces B: Biointerfaces, 62(2), 181–187. https://doi.org/10.1016/j.colsurfb.2007.12.003
CrossRef - Kapoor, D., Bhardwaj, S., Landi, M., & Sharma, A. (2020). The impact of drought stress on secondary metabolites in medicinal and aromatic plants. Plant Physiology and Biochemistry, 156, 437–453. https://doi.org/10.1016/j.plaphy.2020.09.036
CrossRef - Kleinwächter, M., &Selmar, D. (2015). New insights explain that drought stress enhances the quality of spice and medicinal plants: Potential applications. Agronomy for Sustainable Development, 35(1), 121–131. https://doi.org/10.1007/s13593-014-0260-3
CrossRef - Kliebenstein, D. J. (2019). The genetics of plant secondary metabolism. Annual Review of Plant Biology, 70, 285–312. https://doi.org/10.1146/annurev-arplant-050718-100244
- Li, Y., Kong, D., & Wu, H. (2021). Soil pH and ginsenoside profiles in Panax ginsengcultivation: A meta-analysis. Journal of Ginseng Research, 45(4), 567–575. https://doi.org/10.1016/j.jgr.2021.01.003
CrossRef - López-Bucio, J., Cruz-Ramírez, A., & Herrera-Estrella, L. (2018). The role of nutrient availability in regulating root architecture. Current Opinion in Plant Biology, 6(3), 280–287. https://doi.org/10.1016/S1369-5266(03)00035-9
CrossRef - Marschner, H. (2012). Mineral nutrition of higher plants(3rd ed.). Academic Press.
- Maurya, R., & Singh, R. (2020). Effect of soil texture on glycyrrhizin content in Glycyrrhizaglabra roots. Industrial Crops and Products, 145, 112092. https://doi.org/10.1016/j.indcrop.2019.112092
- Mishra, A., &Tanna, B. (2017). Salinity stress and its impact on secondary metabolite production in medicinal plants. Journal of Medicinal Plants Research, 11(41), 645–657. https://doi.org/10.5897/JMPR2017.6460
CrossRef - Ncube, B., Finnie, J. F., & Van Staden, J. (2012). Quality from the field: The impact of environmental factors as quality determinants in medicinal plants. South African Journal of Botany, 82, 11–20. https://doi.org/10.1016/j.sajb.2012.05.009
CrossRef - Omer, E. A., & El-Baroty, G. S. (2020). Influence of nitrogen fertilizer on artemisinin content in Artemisia annua under different water regimes. Egyptian Journal of Agronomy, 42(2), 151–164. https://doi.org/10.21608/agro.2020.104246
- Pott, D. M., & Osorio, S. (2019). The role of potassium in plant secondary metabolism. Journal of Experimental Botany, 70(15), 3835–3847. https://doi.org/10.1093/jxb/erz157
CrossRef - Rengel, Z. (Ed.). (2021). Therhizosphere: An ecological perspective. Academic Press. https://doi.org/10.1016/C2018-0-04434-9
- Sangwan, N. S., Farooqi, A. H. A., & Sangwan, R. S. (2018). Regulation of terpenoid biosynthesis in medicinal plants. In R. S. Sangwan & N. S. Sangwan (Eds.), Biotechnology of medicinal plants(pp. 165–194). Science Publishers.
- Selmar, D., &Kleinwächter, M. (2013). Stress enhances the synthesis of secondary plant products: The impact of stress-related over-reduction on the accumulation of natural products. Plant and Cell Physiology, 54(6), 817–826. https://doi.org/10.1093/pcp/pct054
CrossRef - Sharma, S., & Singh, R. (2020). Heavy metal stress and its effect on phytochemical profiles in HypericumperforatumEnvironmental Science and Pollution Research, 27(21), 26789–26799. https://doi.org/10.1007/s11356-020-09027-5
- Singh, A., & Dubey, N. K. (2019). Influence of soil pH on secondary metabolite production in Catharanthusroseus(L.) G. Don. Journal of Applied Research on Medicinal and Aromatic Plants, 15, 100225. https://doi.org/10.1016/j.jarmap.2019.100225
- Taiz, L., &Zeiger, E. (2015). Plant physiology and development(6th ed.). Sinauer Associates.
- Van Dam, N. M., &Bouwmeester, H. J. (2016). Metabolomics in the rhizosphere: Tapping into belowground chemical communication. Trends in Plant Science, 21(3), 256–265. https://doi.org/10.1016/j.tplants.2016.01.008
CrossRef - Wani, S. H., &Gosal, S. S. (2020). Improving secondary metabolite production in medicinal plants through soil management. Springer. https://doi.org/10.1007/978-981-15-0843-3
- Yang, L., Wen, K. S., Ruan, X., Zhao, Y. X., Wei, F., & Wang, Q. (2018). Response of plant secondary metabolites to environmental factors. Molecules, 23(4), 762. https://doi.org/10.3390/molecules23040762
CrossRef
Accepted on: 03 Mar 2026
Second Review by: Dr. S. K Kumar
Final Approval by: Dr. Tanay Pramanik







ISSN Online: 2231-5039

![]()




{kind=link}